








Néogène




 Muridés
Muridés  Pinnipèdes
Pinnipèdes  Australopithéques
Australopithéques 
 Créodontes
Créodontes Astrapothériens
Astrapothériens Desmostyliens
Desmostyliens




Reconstitution de la tête d'un Australopithèque (photographie, Museum d'histoire naturelle de Bâle).
Résumé
- Orogenèse alpine.
- Refroidissement global; calottes glaciaires; banquise à la fin de la période.
- La mise en communication des continents provoque une intense diversification des faunes de Mammifères. La biodiversité des Mammifères est à son apogée.
- Des super-prédateurs géants dominent les pyramides alimentaires des océans. Les Baleines à fanons sont très diversifiées et les Pinnipèdes font leur apparition.
Néogène
Paléogéographie et climat
Climat
- Température moyenne du globe : 25 à 15 °C.
- Refroidissement global, calottes glaciaires, apparition de la banquise.
- Zones tropicales plus étendues vers le nord qu'à l'époque actuelle.
Atmosphère
- O2 : 21 %
- CO2 : 0.06 à 0.03 %
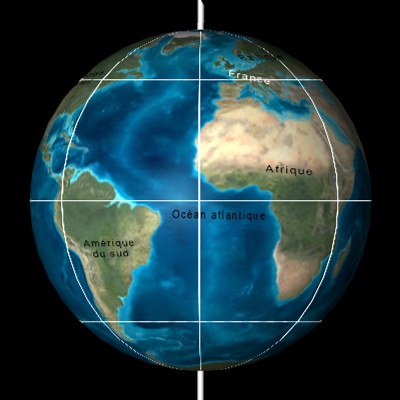
Paléogéographie

Paléogéographie de la France au Miocène. Les fosses molassiques sont des fossés dont le fond s'enfonce progressivement et qui se remplissent des sédiments arrachés aux Alpes, aux Pyrénées et au Massif central. La mer les a envahies à plusieurs reprises. Lorsqu'elle se retirait, des fleuves creusaient des vallées encaissées et chariaient les sédiments.
[10][13][63][28]
L'orogenèse alpine se poursuit et la mer de Téthys se ferme mettant en contact l'Afrique avec le reste du monde. Le Jura se plisse tandis que les fossés d'effondrement du Rhin, du Forez et de la Limagne apparaissent. Un volcanisme intense est associé à ces fossés et la majorité des massifs volcaniques du Massif central (Velay, Cantal, Aubrac, Mont Dore) se forment alors. Fossés et volcanisme sont révélateurs d'un activité de rifting provoquée par l'étirement de la lithosphère européenne lors de sa subduction [64]. Plus au sud, l'activité de rifting provoque la séparation du bloc corso-sarde de la Provence et sa dérive vers l'est. Il atteint sa position actuelle en 7 millions d'années. Une transgression marine envahit la vallée du Rhône jusqu'à Lyon et l'océan Atlantique déborde sur l'Aquitaine et la vallée de la Loire formant alors une mer peu profonde: la Mer des faluns.
A la fin du Miocène, le rapprochement de l'Afrique et de l'Europe ferme le détroit de Gibraltar. La Méditerranée s’assèche et se réduit à quelque lagunes éparses à 1500 m au dessous du niveau actuel. D'importants dépôts de gypse et de sel gemme se forment alors. La réouverture brutale de détroit de Gibraltar permet un remplissage rapide (en quelques mois?) de la Méditerranée [64].
Ailleurs dans le monde, l'Inde continue d'emboutir l'Asie. Les Montagnes rocheuses et la Cordillères des Andes poursuivent leur surrection et leur érosion s'accompagne d'immenses dépôts de sédiments dans l'océon Pacifique et sur les continents. Les sédiments néogènes recèlent d'importantes ressources minérales, champs pétrolifères (Texas, Californie, Louisiane) ou alluvions aurifères de la ruée vers l'or du Far West [63].
La planète continue à se refroidir et le continent Antarctique se couvre de glace. A la fin du Pliocène la banquise apparaît au pôle nord.
La vie sur les continents


Reconstitution d'un paysage du Miocène de la région de Sansan (Gers, France). Un Deinotherium, bien reconnaissable aux défenses recourbées vers le bas de sa mâchoire inférieure, mange les feuilles d'un arbre tandis qu'un autre s'abreuve et que le Chalicothère Anisodon patauge dans l'eau. Un troupeau d'Archaeobelodon traverse le plan d'eau sous le regard attentif de deux Pliopithecus venus boire près du Rhinocéros sans cornes Hoploaceratherium. Au premier plan des Dicrocerus regardent avec inquiétude le tigre à dents de sabre Sansanosmilus s'approcher discrètement caché dans les hautes herbes. L'aigle pêcheur Haliaeetus survole la scène guettant les poissons dérangés par le troupeau d'Archaeobelodon [23] [25] [26] [27] [37] [44]

Le Rhinoceros sans corne Brachypotherium [12] attaqué par des Amphicyon. [5] [22] [52] [53] . A l'arrière plan à gauche des Equidés à 3 doigts Anchitherium et à droite un troupeau d'Archaeobelodon
Au début du Miocène tous les continents sont isolés. Seules l'Europe et l'Asie communiquent depuis la disparition de la mer d'Oural à la fin de l'Oligocène. Chaque continent fonctionne comme un laboratoire de l'évolution indépendant avec son cortège de convergences. Faune et flore ont leur particularité. La faune australienne est exclusivement constituée de Marsupiaux. En amérique du Sud les prédateurs sont uniquement des Marsupiaux et des Oiseaux géants tandis que les végétariens sont des placentaires du groupe des Méridiungulés et des Primates dont l'origine Africaine reste incertaine [58] . Les Afrothériens dominent l'Afrique et l'Amérique du Nord a vu l'évolution des Équidés. Le décor est planté avant le grand chambardement.
Au cours du Néogène, des voies terrestres mettent en contact l'Amérique du Sud avec l'Amérique du Nord (Isthme de panama), Eurasie avec l'Afrique (fermeture de la mer de Téthys et du détroit de Gibraltar). Les migrations qu'elles permettent entraînent d'intenses diversifications dans les espaces nouvellement conquis mais les compétitions qu'elles engendrent provoquent l'extinction de certains groupes (les Méridiungulés, les Marsupiaux carnivores et les Oiseaux géants en Amérique du sud [19] [46] [59], la plupart des espèces d'Hyracoïdes en Afrique) [48] [56] . De ces guerres d'invasion, les Mammifères nord-américains et eurasiatiques sortent vainqueur. Une exception notable: celle des Proboscidiens (actuels Éléphants) qui vont tirer un net avantage de leur colonisation de l'Eurasie et des Amériques. Les Primates africains sont aussi du voyage et envahissent l'Eurasie. Mais les faits marquants de leur diversification se produisent en Afrique avec, par exemple, l'apparition des Australopithèques il y a 4 millions d'années.
La paléogéographie n'est pas le seul facteur de diversification. Le refroidissement et l'assèchement progressif du climat global de la planète entraîne la zonation de la végétation et l'apparition d'une multitude de milieux divers. A cela s'ajoute le développement des monocotylédones herbacées et la multiplication des prairies, steppes et autres étendues herbeuses. La chaîne alimentaire herbe ——> herbivore ——> carnivore est alors pleinement efficace permettant l'augmentation de la taille des animaux. Au miocène, la diversité des Mammifères est à son maximum.
La vie dans les océans


Au Miocène vous auriez pu déguster des huîtres géantes de 30 cm (Ostrea crassissima) et récolter des Murex, des Carditas, des Pectens mais aussi des Oursins et des Nautiles dans les mers peu profondes et des Cérithes dans les lagunes. A cette faune d'invertébrés, il faut ajouter des Siréniens, des Baleines et les derniers arrivés dans les océans: les Pinnipèdes (Phoques, Lions de mer et Otaries). Dans les cours d'eau et les lacs la pêche aurait été fructueuse: Tanches, Perches, Gardons et Anguilles côtoyaient des espèces des régions tropicales.
Nous ne saurions passer sous silence deux super-prédateurs célèbres: le Mégalodon et le Livyatan.

La première reconstitution de la mâchoire de Megalodon exposée à l'American Museum of Natural History de New-York en 1909 [17]. Seules sont réelles les dents, le reste a été recréé à partir des os de la mâchoire du Grand Requin Blanc. La largeur de la mâchoire est exagérée.
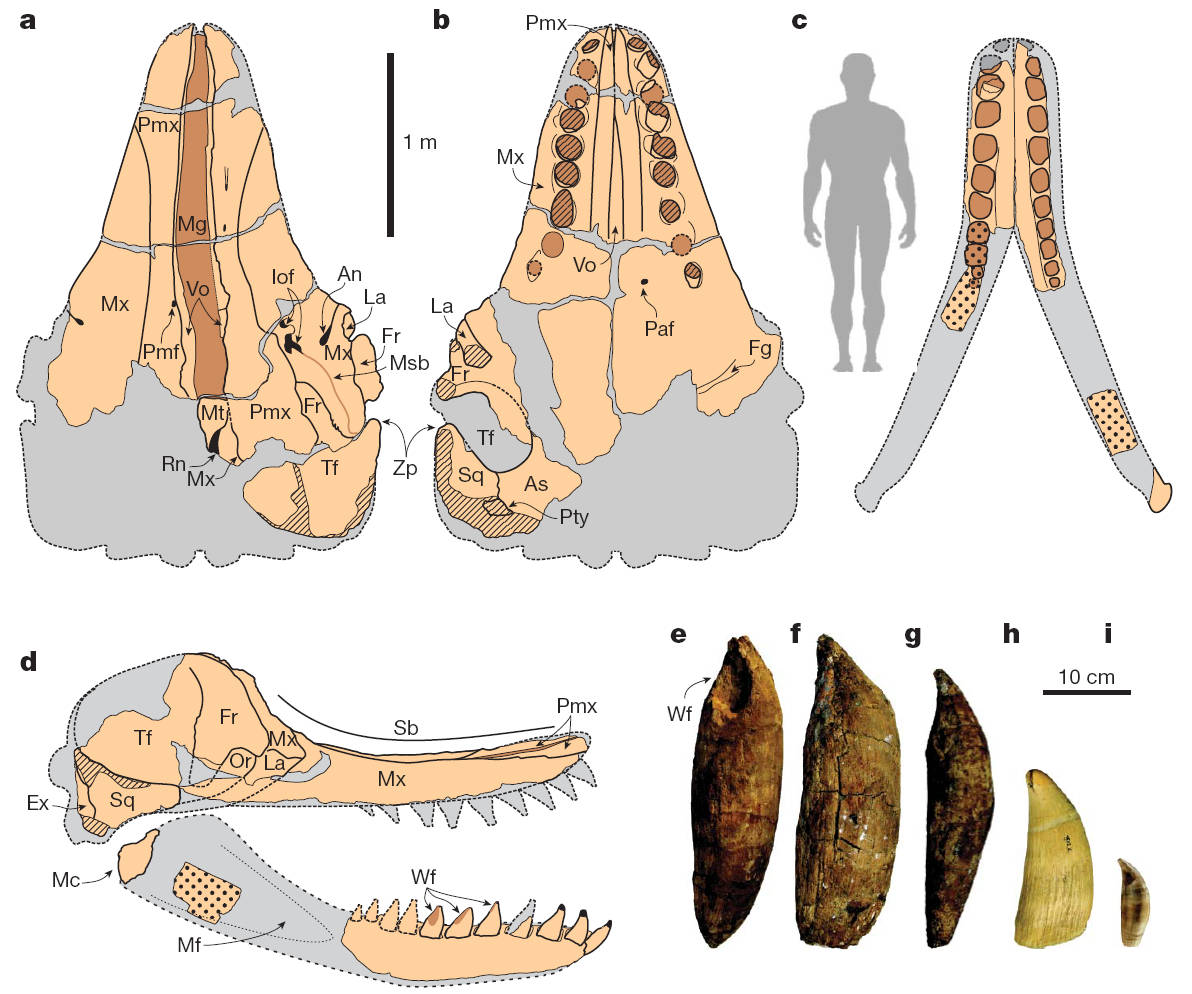
Crâne en vue dorsale (a), en vue ventrale (b); mandibule en vue dorsale (c); crâne et mandibule en vue latérale (d) de Livyatan melvillei. Comparaison des dents de Livyatan melvillei (e, f, g) avec celles du Cachalot (h) et de l'Orque (i)[36].
Le Mégalodon (Otodon megalodon) était un Requin géant pouvant atteindre 20 m de long soit deux ou trois fois la longueur du Grand Requin Blanc actuel (Carcharodon carcharias). Cette longueur est une estimation car on n'a jamais trouvé de squelette complet de l'animal mais seulement quelques morceaux de crâne, des séries de vertèbres et surtout des dents dont les plus grandes pouvaient atteindre 16 cm.
Le Livyatan (Livyatan melvillei) était un Cachalot dont la longueur estimée est de 13 à 17 mètres, là encore on ne connaît pas de squelette complet, seuls le crâne et la mandibule nous sont parvenus. Le crâne mesure 3 mètres de long et il est armé, tout comme la mandibule, d'une batterie de dents impressionnantes de plus de 30 cm. Contrairement aux Cachalots actuels qui gobent des Calmars géants, le Livyatan dévorait des Baleines [36].

Les Requins géants et le Megalodon ont fait fantasmer les foules. Hollywood en a fait des monstres sacrés avec les "Dents de la mer" et plus récemment "En eaux troubles". Même si Megalodon mesurait 20 mètres ce n'était pas le plus grand Requin de tout les temps. Le Requin baleine actuel peut lui aussi atteindre 20 mètres de long mais il est vrai qu'il est bien moins cinégénique dans le rôle d'un placide mangeur de plancton. Livyatan, aussi impressionnant qu'il soit, entre dans la gamme de taille des Cachalots actuels. Mais ces deux "monstres" n'aurait fait qu'une bouchée de Jason Statham et c'est leur régime alimentaire qui interpelle le biologiste. Il fallait des proies de tailles conséquentes pour rassasier ces animaux. Est-ce un hasard s'ils ont vécu à l'âge d'or des Baleines à fanons lorsque la diversité de ces dernières était 5 fois plus élevée qu'à l'heure actuelle?
"Jaws", "Les dents de la mer" en français, met en scène un Requin géant qui est un nain par rapport au Megalodon surdimensionné et bodybuildé de "En eaux troubles".
Sites fossilifères remarquables
| MA | SERIES | ETAGES | Nom et localisation | Fossiles | Milieu |
|---|---|---|---|---|---|
|
-2,58 -3,6 |
Pliocène | Piacenzien | Sterkfontein, Afrique du sud [64] | Chacals, Antilopes, Australopithecus africanus, Baboins, Felins (Homotherium). | Savane arborée. |
| -5,33 | Zancléen | Laetoli Tanzanie [64] | Australopithecus, Eléphants (Loxodonta), Girafe (Giraffa), Rhinoceros (Diceros), Pintades (Numida). | Savane arborée près d'un lac. | |
| -7,25 | Miocène | Messinien | Désert de Djurab Tchad [64] | Sahelanthropus, Artiodactyles (Sivatherium), Périssodactyles (Hipparion, Kobus), Félins (Machairodus), Hippopotames (Hexoprotodon), Hyènes (Ictitherium). | Bord d'un Lac. |
| --11,63 | Tortonien | Saint-Bauzile, Ardèche, France [41] | Squelettes entiers (restes charboneux de la peau et de la chair, trace de la fourrure), feuilles, Invertébrés, Reptiles, Batraciens, Oiseaux, Mammifères (Mulots, Lièvres, Sangliers, Antilopes, Rhinocéros, Chevaux). | Lac à diatomées prés d'un volcan. | |
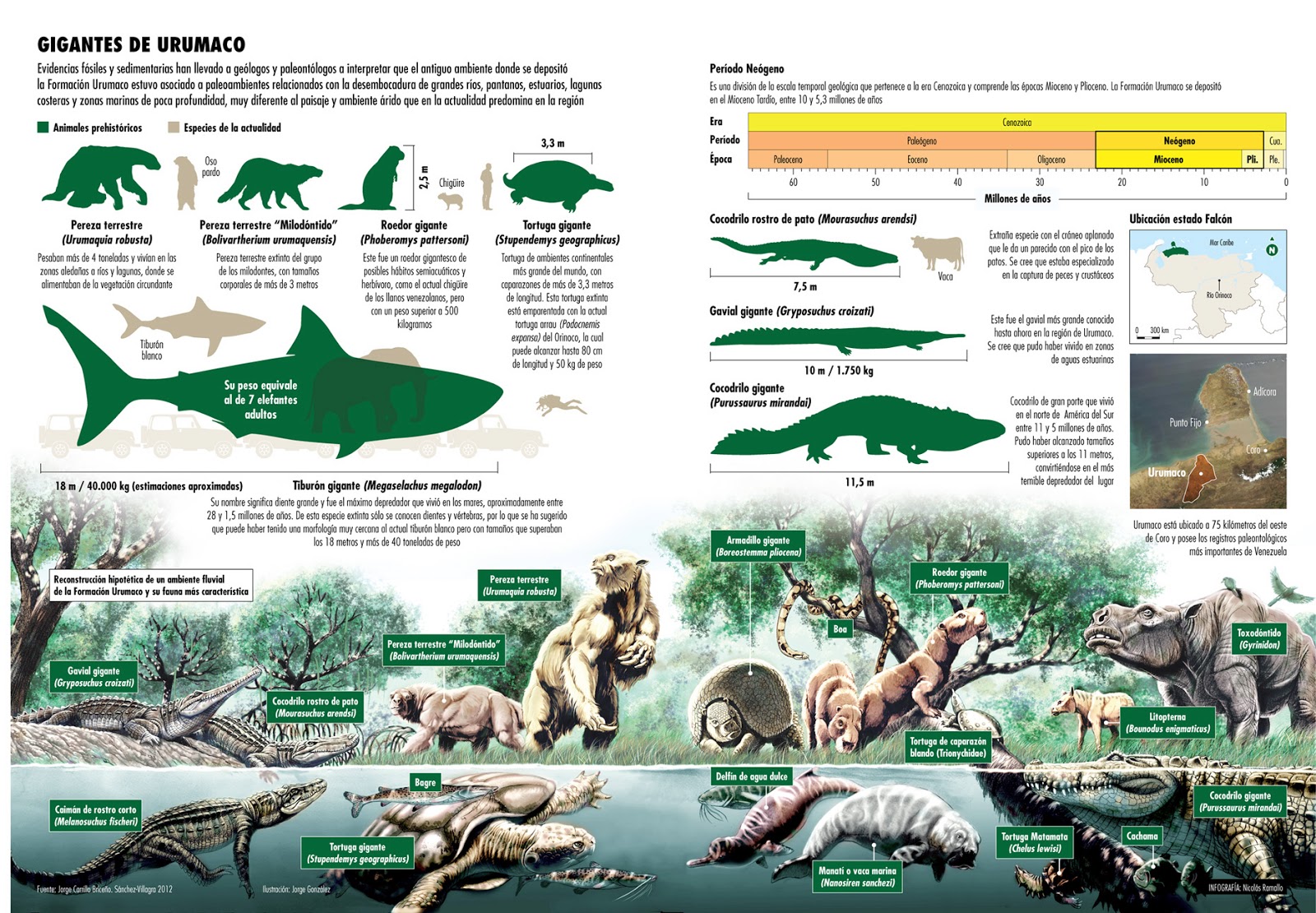
| Urumaco Formation, Venezuela [47] | Faune riche en Reptiles (Tortues et Crocodiles géants), Paresseux géants (Urumaquia), Litopternes, Toxodontes, Dauphins d'eau douce (Ischyrorhynchus), Siréniens (Nanosiren). | Rivières, marécages, savanes, océan. | |||
| -13,82 | Serravallien | Siwalik Hills, Inde et Pakistan [64] [66] [67] | Singes (Sivapithecus), Chalicotherium, Proboscidiens (Gomphotherium, Platybelodon), Artiodactyles (Protragocerus, Giraffokeryx), prédateurs (Percrocuta, Hyainailurus). | Terrestre et fleuve. | |
| -15,97 | Langhian |
Sansan, Gers, France [27] [44] |
Cervidés (Dicrocerus), Proboscidiens (Deinotherium, Archarobelodon), Rhinoceros (Hoploaceratherium), Primates (Pliopithecus)Prédateurs (Sansanosmilus, Amphicyon) Oiseaux et Reptiles (Tortues et Crocodiles). | Forêt au bord d'un plan d'eau. | |
| -20,44 | Burdigalien | Santa Cruz Formation, Argentine, Chili [65] | Oiseaux géants (Phorusrhacos) Meridiungulés, Paresseux terrestres, Tatous, Marsupiaux carnivores (Borhyaena, Lycopsis), rongeurs, Primates (Homunculus). | Savanes, forêts galerie, forêts, cours d'eau. | |
| Rusinga Island, Kenya [64] | Nombreux Singes dont le (Proconsul), Proboscidiens (Gomphotherium), Chalicotherium, prédateur (Créodonte Hyainailouros). | Forêt au bord d'un lac. | |||
| -23.03 | Aquitanien | Riversleigh, Queensland, Australie [64] | Oiseaux géants ( Bullockornis) et Marsupiaux, gros herbivores (Namilamadeta, Neohelos), Sarrigues, Koala (Litokoala), Prédateurs (Priscileo et Kangourou carnivore Ekaltadeta). | Lac bordé d'une forêt pluviale. |
Bibliographie (69 références)
Agustí J. - 2018. Las faunas de mamíferos del Mioceno continental de la Península Ibérica. revista PH:182.
Anquetin J., Antoine P. O., and Tassy P. - 2007. Middle Miocene Chalicotheriinae (Mammalia, Perissodactyla) from France, with a discussion on chalicotheriine phylogeny. Zoological Journal of the Linnean Society 151(3):577–608.
Anton M. - 2003. Reconstructing fossil mammals: strengths and limitations of a methodology. Palaeontological Association Newsletter 53:55–65.
Antón M., Siliceo G., Pastor J. F., and Salesa M. J. - 2022. Concealed weapons: A revised reconstruction of the facial anatomy and life appearance of the sabre-toothed cat Homotherium latidens (Felidae, Machairodontinae). Quaternary Science Reviews 284:107471.
Argot C. - 2010. Morphofunctional analysis of the postcranium of Amphicyon major (Mammalia, Carnivora, Amphicyonidae) from the Miocene of Sansan (Gers, France) compared to three extant carnivores: Ursus arctos, Panthera leo, and Canis lupus. Geodiversitas 32(1):65–106.
Arias-Martorell J., Alba D. M., Potau J. M., Bello-Hellegouarch G., and Pérez-Pérez A. - 2015. Morphological affinities of the proximal humerus of Epipliopithecus vindobonensis and Pliopithecus antiquus: Suspensory inferences based on a 3D geometric morphometrics approach. Journal of Human Evolution 80:83–95.
Azanza B., DeMiguel D., and Andrés M. - 2011. The antler-like appendages of the primitive deer Dicrocerus elegans : morphology, growth cycle, ontogeny, and sexual dimorphism. Estudios Geologicos 67(2):579–602.
Bai B., Wang Y. Q., and Meng J. - 2018. The divergence and dispersal of early perissodactyls as evidenced by early Eocene equids from Asia. Communications Biology 2018 1:1 1(1):1–10.
Berta A., Ray C. E., and Wyss A. R. - 1989. Skeleton of the Oldest Known Pinniped, Enaliarctos mealsi. Science (New York, N.Y.) 244(4900):60–62.
Besson D. - 2005. Architecture du bassin rhodano-provençal miocène (Alpes, SE France) : relations entre déformation, physiographie et sédimentation dans un bassin molassique d’avant-pays. Thèse de Sciences de la Terre. École Nationale Supérieure des Mines de Paris.
Bucknall E., Woodward A. B., and Knipe H. R. - 1912. Evolution in the past. (Herbert and Daniel, Ed.), London.
Cerdeno E. - 1993. Etude sur Diaceratherium aurelianense et Brachypotherium brachypus (Rhinocerotidae, Mammalia) du Miocene moyen de France. Bulletin - Museum National d’Histoire Naturelle Section C: Sciences de la Terre 15(1–4):25–77.
Chaix C., and Cahuzac B. - 2005. Les faunes de Scléractiniaires dans les faluns du Miocène moyen d’Atlantique-Est (bassins de la Loire et d’Aquitaine) : paléobiogéographie et évolution climatique. Annales de Paléontologie 91(1):33–72.
Coombs M. - 1989. Interrelationships and diversity in the Chalicotheriidae.in D. R. Prothero and R. M. Schoch, editor. The Evolution of Perissodactyls. Clarendon Press/Oxford University Press.
Coombs M. C., and Cote S. M. - 2012. Chalicotheriidae. Cenozoic Mammals of Africa:659–668.
Cuvier G., and Laurillard M. - 1850. Anatomie comparée, planches de myologie.(Dusacq, Ed.).
Dean. B. - 1909. The giant of ancient sharks. The American Museum Journal 9(8):232–234.
Depéret C. - 1892. La faune de mammifères miocènes de la Grive-Saint-Alban (Isère) et de quelques autres localités du bassin du Rhône. — Documents nouveaux et révision générale. Publications du musée des Confluences 5(1):1–15.
Domingo L., Tomassini R. L., Montalvo C. I., Sanz-Pérez D., and Alberdi M. T. - 2020. The Great American Biotic Interchange revisited: a new perspective from the stable isotope record of Argentine Pampas fossil mammals. Scientific Reports 2020 10:1 10(1):1–10.
Elmi S., Babin C., and Grosheny D. - 2020. Histoire de la Terre. Dunod, Ed..
Etienne C., Houssaye A., and Hutchinson J. R. - 2021. Limb myology and muscle architecture of the Indian rhinoceros Rhinoceros unicornis and the white rhinoceros Ceratotherium simum (Mammalia: Rhinocerotidae). PeerJ 9.
Fahlke J. M., Coombs M. C., and Semprebon G. M. - 2013. Anisodon sp. (Mammalia, Perissodactyla, Chalicotheriidae) from the Turolian of Dorn-Dürkheim 1 (Rheinhessen, Germany): Morphology, phylogeny, and palaeoecology of the latest chalicothere in Central Europe. Palaeobiodiversity and Palaeoenvironments 93(2):151–170.
Figueirido B., Perez-Claros J. A., Hunt R. M. J., and Palmqvist P. - 2011. Body mass estimation in amphicyonid carnivoran mammals: A multiple regression approach from the skull and skeleton. Acta Palaeontologica Polonica 56(2).
Fihlhol H. - 1891. Études sur les mammifères fossiles de Sansan. Masson.
Flynn L. J., Morgan M. E., Pilbeam D., and Barry J. C. - 2014. “endemism” Relative to space, time, and taxonomic level. Annales Zoologici Fennici 51(1–2):245–258,
Flynn L. J., Lindsay E. H., Pilbeam D., Raza S. M., Morgan M. E., Barry J. C., Badgley C. E., Behrensmeyer A. K., Cheema I. U., Rajpar A. R., and Opdyke N. D. - 2015. The Siwaliks and Neogene Evolutionary Biology in South Asia. Fossil Mammals of Asia:353–372,
Frick C. - 1933. New remains of trilophodont-tetrabelodont mastodons. Bulletin of the AMNH ; v. 59, article 9..
Gervais P. - 1859. Zoologie et paléontologie françaises : nouvelles recherches sur les animaux vertébrés dont on trouve les ossements enfouis dans le sol de la France et sur leur comparaison avec les espèces propres aux autres régions du globe.
Ginsburg L. - 1961. La faune des Carnivores miocènes de Sansan. Mémoires du Muséum national d'histoire naturelle, NS, sér. C, IX : 1-190, 72 fig., 20 pl.
Ginsburg L. - 2000. La faune miocene de sansan et son environnement. Mémoires du Muséum national d’histoire naturelle 183(1):392.
Ginsburg L., and Al. - 1998. Le Miocène du bassin de Vence (Alpes-Maritimes, France) : stratigraphie et paléogéographie. Geodiversitas 20(2):229–238.
Göhlich U. B. - 2010. Tertiäre Urelefantenfunde aus Deutschland. Pages 363–372 in H. Meller, editor. Elefantenreich – Eine Fossilwelt in Europa. Elefantenr. Landesamt für Denkmalpflege und Archäologie Sachsen-Anhalt.
Gratiolet P.-L. (1815-1865). A. du texte. - 1867. Recherches sur l’anatomie de l’hippopotame / par Louis-Pierre Gratiolet; publiées par les soins du Dr Edmond Alix:406.
Gregor H. J., and Fiest W. - 1988. Das Biotop und die Nahrung des Molasse-Affen Pliopithecus antiquus. documenta naturae 42:28–36.
Harrison T. - 2013. Catarrhine origins. Pages 376–396 A Companion to Paleoanthropology. Blackwell Publishing Ltd..
Hocking D. P., Marx F. G., Sattler R., Harris R. N., Pollock T. I., Sorrell K. J., Fitzgerald E. M. G., McCurry M. R., and Evans A. R. - 2018. Clawed forelimbs allow northern seals to eat like their ancient ancestors. Royal Society Open Science 5(4).
Holland, W.J.; Peterson O. A., - 1913. The osteology of the Chalicotheroidea. With special reference to a mounted skeleton of Moropus elatus Marsh, now installed in the Carnegie museum. Pittsburgh Memoirs Carnegie Museum 3(2):189–411.
Kirk E. C., and Simons E. L. - 2001. Diets fo fossil primates from the fayum depression of Egypt: A quantitative analysis of molar shearing. Journal of Human Evolution 40(3):203–229.
Lambert O., Bianucci G., Post K., De Muizon C., Salas-Gismondi R., Urbina M., and Reumer J. - 2010. The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru. Nature 2010 466:7302 466(7302):105–108.
Lartet É. - 1851. Notice sur la colline de Sansan, suivie d’une récapitulation des diverses espèces d’animaux vertébrés fossiles, trouvés soit à Sansan, soit dans d’autres gisements du terrain tertiaire du Miocène dans le bassin sous-pyrénéen par Ed. Lartet, M. Noulet, M. l’Abbé Dupuy, M. de Boissy,
Lesbre F.-X. (1858-1942). A. du texte. - 1897. Essai de myologie comparée de l’homme et des mammifères domestiques, en vue d’établir une nomenclature unique et rationnelle, par M. F.-X. Lesbre.Société d’anthropologie de Lyon.
MacLaren J. A., and McHorse B. K. - 2020. Comparative forelimb myology and muscular architecture of a juvenile Malayan tapir (Tapirus indicus). Journal of Anatomy 236(1):85–97.
MacLaren J. A., and Nauwelaerts S. - 2020. Modern Tapirs as Morphofunctional Analogues for Locomotion in Endemic Eocene European Perissodactyls. Journal of Mammalian Evolution 27(2):245–263.
Métais G., and Sen S. - 2018. The late Miocene mammals from the Konservat-Lagerstätte of Saint-Bauzile (Ardèche, France). Comptes Rendus Palevol 17(7):479–493.
Morales J., Sánchez I., and Quiralte V. - 2012. Les Tragulidae (artiodactyla) de Sansan. Mémoires du Muséum national d´Histoire naturelle 203:225–247.
Morlo M., Peigné S., and Nagel D. - 2004. A new species of Prosansanosmilus: implications for the systematic relationships of the family Barbourofelidae new rank (Carnivora, Mammalia). Zoological Journal of the Linnean Society 140(1):43–61.
Palmer D., Barrett P., and Leibovici A. - 2009. L'évolution l'histoire de la vie. Guy Trénadiel éditeur 374 pp.
Peigné S., and Sen S. - 2012. Mammifères de Sansan. 709p. Mémoires du Muséum national d'Histoire naturelle ; 203.
Peigné S., Salesa M. J., Antón M., and Morales J. - 2008. A new amphicyonine (carnivora: amphicyonidae) from the upper miocene of batallones-1, madrid, spain. Palaeontology 51(4):943–965.
Pomerol Ch. - 1973. Stratigraphie et paléogéographie, Ère cénozoïque: tertiaire et quaternaire. Douin éditeur 269p
Prevosti F. J., Forasiepi A., and Zimicz N. - 2013. The Evolution of the Cenozoic Terrestrial Mammalian Predator Guild in South America: Competition or Replacement? Journal of Mammalian Evolution 20(1):3–21.
Sánchez-Villagra M. R., and Aguilera O. A. - 2006. Neogene vertebrates from Urumaco, Falcón State, Venezuela: Diversity and significance. Journal of Systematic Palaeontology 4(3):213–220.
Sanmartín I., Enghoff H., and Ronquist F. - 2001. Patterns of animal dispersal, vicariance and diversification in the Holarctic. Biological Journal of the Linnean Society 73(4):345–390.
Schötz G. M. - 2005. Ein Skelettrest von Archaeobelodon aus der Oberen Süßwassermolasse (Miozän) von Junkenhofen bei Gerolsbach (Krs. Pfaffenhofen a.d. Ilm). Berichte des Naturwissenschaftlichen Vereins für Schwaben 109:5–25.
Schultz C., Schultz M., and Martin L. - 1970. A New Tribe of Saber-toothed Cats(Barbourofelini) from the Pliocene of North America. Bulletin of the University of Nebraska State Museum. 9(1):1–31.
Semprebon G. M., Pirlo J., and Dudek J. - 2022. Dietary Habits and Tusk Usage of Shovel-Tusked Gomphotheres from Florida: Evidence from Stereoscopic Wear of Molars and Upper and Lower Tusks. Biology 11(12).
Siliceo G., Salesa M. J., Antón M., Peigné S., and Morales J. - 2017. Functional anatomy of the cervical region in the late Miocene amphicyonid Magericyon anceps (Carnivora, Amphicyonidae): implications for its feeding behaviour. Palaeontology 60(3):329–347.
Sorkin B. - 2006. Ecomorphology of the giant bear-dogs Amphicyon and Ischyrocyon. Historical Biology 18(4):375–388.
Tassy P. - 2013. L’anatomie cranio-mandibulaire de Gomphotherium angustidens (Cuvier, 1817) (Proboscidea, Mammalia): Données issues du gisement d’En Péjouan (Miocène moyen du Gers, France). Geodiversitas 35(2):377–445.
Tassy P. - 2014. L’odontologie de Gomphotherium angustidens (Cuvier, 1817) (Proboscidea, Mammalia) : données issues du gisement d’En Péjouan (Miocène moyen du Gers, France). Geodiversitas 36(1):35–115.
Turner A., Antón M., Salesa M. J., and Morales J. - 2012. Changing ideas about the evolution and functional morphology of Machairodontine felids. Estudios Geologicos 67(2):255–276.
Wen J., Nie Z. L., and Ickert-Bond S. M. - 2016. Intercontinental disjunctions between eastern Asia and western North America in vascular plants highlight the biogeographic importance of the Bering land bridge from late Cretaceous to Neogene. Journal of Systematics and Evolution 54(5):469–490.
Werdelin L., Yamaguchi N., Johnson W., and O’Brien S. J. . - 2010. Phylogeny and evolution of cats (Felidae). Pages 59–81 in A. J. L. D.W. Macdonald, editor. Biology and Conservation of Wild Felids. Oxford University Press.
« Santa Cruz Formation » (2024, 15 janvier). Dans Wikipedia https://en.wikipedia.org/wiki/Santa_Cruz_Formation consulté le: 2 avril 2024
Wisniewski A. L., Lloyd G. T., and Slater G. J. - 2022. Extant species fail to estimate ancestral geographical ranges at older nodes in primate phylogeny. Proceedings of the Royal Society B 289(1975).
Woodburne M. O. - 2010. The Great American Biotic Interchange: Dispersals, Tectonics, Climate, Sea Level and Holding Pens. Journal of Mammalian Evolution 17(4):245.
Yu W., Jin M., PalAsiatica Y. J.-V., and 2003 undefined. (n.d.). The discovery of pliopithecus from northern junggar basin, xinjiang. vertpala.ac.cnWUW Yu, M Jin, Y JieVertebrata PalAsiatica, 2003•vertpala.ac.cn.
Zapfe H. - 1976. Die Fauna der miozänen Spaltenfüllung von Neudorf a. d. March (ČSSR.) Chalicotherium grande. Sitzungsberichte der Akademie der Wissenschaften mathematisch-naturwissenschaftliche Klasse 185:91–112.
Zapfe H. - 1979. Chalicotherium grande (Blainv.) aus der miozδnen Spaltenfόllung von Neudorf an der March (Devvinskα Novα Ves),Tschechoslwakei. Neue Denkschriften des Naturhistorischen Museums in Wien 2:1–282.





















{kind=link}